Translácia predstavuje konečný proces realizácie genetickej informácie štruktúrnych génov. Prebieha na ribozómoch v cytoplazme alebo membránach endoplazmatického retikula za účasti transferových RNA (tRNA), ktoré do ribozómov prinášajú jednotlivé aminokyseliny. Na ribozómoch, ktoré sú akýmisi miniatúrnymi továrňami na výrobu bielkovín, prebieha polymerizácia aminokyselín do súvislého polypeptidového reťazca na základe genetického kódu.
Niekedy sa pod pojmom proteosyntéza rozumie translácia aj transkripcia DNA do mRNA. Nemalo by tomu tak byť, pretože transkripcia všeobecne zahŕňa prepis nielen štruktúrnych génov. Odporúčam preto pojem proteosyntéza používať len pre transláciu (napokon je jasné, že každej translácii predchádza transkripcia mRNA).
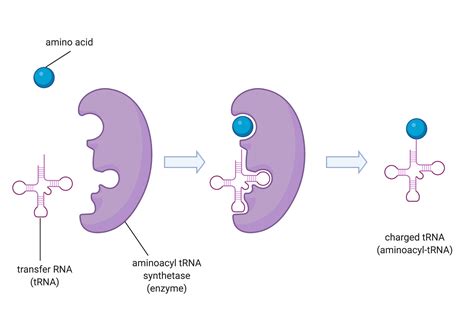
Transferové RNA sú typom funkčných RNA, ktoré slúžia na prepravu, alebo transfer (odtiaľ ich názov), jednotlivých aminokyselín do ribozómu, kde sa z nich tvorí súvislý polypeptidový reťazec. Každá tRNA predstavuje jednovláknovú RNA dlhú 75−93 báz, ktorá okrem typických RNA nukleotidov obsahuje aj neobvyklé nukleotidy.
Štruktúra tRNA
Sekundárna štruktúra tRNA pripomína „ďatelinový štvorlístok”. Jednotlivé „lístky” sú zložené z tzv. palindrómov. Palindrómy sú také nukleotidové sekvencie (RNA alebo DNA), ktoré sú symetrické tak, že bázy od osi symetrie smerom doľava sú komplementárne k bázam nachádzajúcim sa vpravo od osi symetrie, napr. 5'-CAGT|ACTG-3'. To spôsobuje na jednovláknovej molekule tvorbu vnútromolekulových spojení, ktoré lokálne tvoria dvojvláknový úsek nazývaný vlásenka (angl. hairpin).
Ak sa v strede palindrómu, t.j. v oblasti stredu symetrie nachádzajú nukleotidy, ktoré sa nevedia takýmto spôsobom popárovať, na konci vlásenky sa tvorí jednovláknová slučka (angl. loop). Takýmto prípadom vlásenky so slučkou (angl. hairpin loop) sú aj „lístky” tRNA, ktoré sa označujú ako ramená:
- akceptorové rameno - na jeho koniec sa kovalentne viaže aminokyselina
- antikodónové rameno s antikodónovou slučkou - týmto koncom tRNA rozoznáva genetický kód „zapísaný” v mRNA
- pseudouridínové rameno s TΨC-slučkou - obsahuje modifikovaný uracil − pseudouracil
- dihydrouridínové rameno s D-slučkou - obsahuje modifikovaný uracil − dihydrouracil
- variabilná slučka - môže byť krátka (tRNA typu I) alebo dlhá (tRNA typu II), neovplyvňuje biologickú funkciu tRNA
Obidva konce RNA (3' aj 5') sa nachádzajú v akceptorovom ramene tRNA. Na každú tRNA sa pripája len jedna aminokyselina, a to na posledný adenín 3'-konca tRNA, ktorá končí jednovláknovou prečnievajúcou sekvenciou 5'-CCA-3'.
Pre každú z 20 proteínogénnych (tvoriacich proteíny) aminokyselín existuje osobitný enzým − aminoacyl-tRNA-syntetáza, ktorý vytvára kovalentnú väzbu medzi tRNA a jej príslušnou aminokyselinou. Rovnaké aminokyseliny môžu byť spájané s rôznymi tRNA, ale rovnaké tRNA viažu len jednu špecifickú aminokyselinu, čo súvisí s degenerovanosťou genetického kódu.
Všetky tRNA nadobúdajú v živom systéme podobnú terciárnu štruktúru tvaru „Γ”, ktorou najlepšie pasujú do vstupných miest na ribozóme.
Štruktúra tRNA
Genetický kód a kodóny
Zaradenie aminokyseliny do polypeptidového reťazca je dané pomocou troch za sebou idúcich nukleotidov. Takáto trojica nukleotidov, ktorá kóduje určitú aminokyselinu, sa nazýva kodón. Genetický kód je teda systém pravidiel, podľa ktorých jednotlivé kodóny určujú zaradenie štandardných aminokyselín do polypeptidového reťazca. Kodónmi sa označujú sekvencie nukleotidov mRNA, ku ktorým sú komplementárne antikodóny na antikodónovom ramene tRNA. Čítanie genetického kódu teda znamená rozpoznávanie kodónov na mRNA pomocou antikodónov na tRNA a prebieha na ribozóme počas proteosyntézy. Prebieha presne od jedného kodónu k druhému, čo znamená, že jednotlivé trojice nukleotidov sa pri čítaní neprekrývajú.
Niekedy sa zvykne genetický kód v literatúre alebo softvéri interpretovať už na úrovni DNA, a vtedy sa najčastejšie hovorí len o „tripletoch”. Označenie „kodón” je v súvislosti s DNA nepresné. Taktiež si treba dávať pozor na orientáciu DNA, pretože po správnosti by sa mala udávať sekvencia 3',5' reťazca, pretože len toto vlákno DNA je kódujúce a je k nemu komplementárna mRNA.
Uvediem známy príklad so štart kodónom AUG, ktorý kóduje zaradenie metionínu: 5'-AUG-3' je kodón mRNA, ktorému je komplementárny triplet 3'-TAC-5' na DNA, ale zároveň k tomuto tripletu je komplementárny 5'-ATG-3'. Keďže ale vlákno 5',3' na DNA nie je kódujúce (nie je predmetom prepisu RNA-polymerázou), nemali by sme tvrdiť, že triplet ATG kóduje metionín.
Genetický kód pozostáva zo 64 kodónov (4³ = 64). Zmysel kodónu je jeho schopnosť kódovať aminokyselinu. U drvivej väčšiny živých organizmov má genetický kód rovnaký zmysel. Táto vlastnosť genetického kódu sa nazýva univerzálnosť. Od univerzálneho genetického kódu existujú len minimálne odchýlky (u niektorých nižších skupín organizmov a u mitochondrií).
Niektoré kodóny nekódujú žiadnu aminokyselinu, ale majú signalizačnú funkciu. Niekedy sa nazývajú nezmyselné kodóny. Signalizácia je potrebná na vymedzenenie sekvencie mRNA, ktorá bezprostredne súvisí so syntézou proteínov, teda určuje začiatok a koniec syntézy polypeptidového reťazca (pretože nie celá mRNA je prekladaná do sekvencie aminokyselín). Takýto úsek na mRNA, ktorý obsahuje úplnú informáciu pre syntézu polypeptidu, sa nazýva otvorený čítací rámec (angl. open reading frame − ORF).
Začiatok syntézy je určený iniciačným kodónom AUG, ktorý kóduje aminokyselinu metionín (okrem signalizačnej funkcie má teda aj zmysel). Koniec syntézy polypeptidu signalizuje jeden z terminačných kodónov UAA, UAG alebo UGA, ktoré nekódujú žiadnu aminokyselinu.
Čiže kodónov, ktoré kódujú aminokyseliny, je dohromady 61 (64 bez 3 terminačných kodónov). Proteinogénnych aminokyselín je však len 20. Z toho vyplýva, že niektoré aminokyseliny (nie všetky ale väčšina) sú kódované rôznymi kodónmi (synonymné kodóny). To je umožnené existenciou rôznych tRNA, ktoré majú odlišné antikodóny ale nesú rovnakú aminokyselinu. Táto vlastnosť genetického kódu sa nazýva degenerovanosť.
Genetický kód
V tabuľke genetického kódu sa kodóny, ktoré kódujú tú istú aminokyselinu, odlišujú hlavne v tretej pozícii nukleotidu. Ak sa zmenou 3. nukleotidu v kodóne nezmení jeho zmysel, takáto štvorica kodónov sa nazýva kodónová rodina. Dva synonymné kodóny, ktoré majú na 3.

Tabuľka genetického kódu
Ribozómy
Ribozómy tvoria súčasť každej prokaryotickej a eukaryotickej bunky. Ich úlohou je syntéza proteínov v procese translácie. V priemere majú ~20 nm a skladajú sa zo 65 % ribozomálnej RNA (rRNA) a 35 % ribozomálnych proteínov. Predstavujú teda komplex ribonukleovej kyseliny a proteínov (ribonukleoproteínový komplex). Ribozómy boli prvýkrát pozorované špeciálnou technikou mikroskopovania v tmavom poli v 30-ich rokoch 20. storočia a neskôr elektrónovým mikroskopom, kedy bola bližšie charakterizovaná ich štruktúra.
Ribozómy nie sú organely, pretože nie sú obalené žiadnou membránou od okolitej cytoplazmy.
Prokaryotické a eukaryotické ribozómy sa odlišujú vo veľkosti a zložení. Často používaným meradlom hmotnosti ribozómov je sedimentačný koeficient. Udáva sa v jednotkách Svedberg (S), po nositeľovi Nobelovej ceny za chémiu (1926) THEODOROVI SVEDBERGOVI (1884−1971), ktoré určujú rýchlosť sedimentácie častíc pri ultracentrifugácii.
Voľné ribozómy sú lokalizované v cytoplazme prokaryotickej a eukaryotickej bunky, kde syntetizujú proteíny. Niektoré proteíny však vyžadujú, aby ich syntéza bola oddelená od cytoplazmy. Patria k nim napr. proteíny bohaté na cysteíny, ktoré tvoria disulfidické väzby, zložené proteíny (napr. glykoproteíny) alebo proteíny slúžiace na sekréciu z bunky. Takéto proteíny veľmi krátko po začiatku svojej syntézy nasmerujú voľné ribozómy na membránu endoplazmatického retikula (ďalej ER), kde už ich syntéza pokračuje do cisterien ER.
Ribozómy pozostávajú z malej a veľkej ribozomálnej podjednotky. U eukaryot dochádza k ich formovaniu už v jadre, odkiaľ sa hotové podjednotky dostávajú cez jadrové póry do cytoplazmy.
Na ribozóme sú definované viaceré miesta, ktoré interagujú s jednotlivými molekulami (mRNA, tRNA, pomocné proteíny) zúčastňujúcimi sa procesov translácie:
- väzbové miesto pre mRNA - umiestnené na malej podjednotke
- aminoacylové miesto (A miesto) - vstupujú sem tRNA, na ktorých je naviazaná príslušná aminokyselina
- peptidylové miesto (P miesto) - v tomto mieste nastáva predlžovanie polypeptidového reťazca práve o jednu aminokyselinu
- peptidyltransferázové miesto - tvorba peptidovej väzby
- výstupné miesto pre deacylovanú tRNA (E miesto) - tRNA, ktorá odovzdá svoju aminokyselinu, odchádza z ribozómu v tomto mieste
- väzbové miesta pre pomocné translačné proteíny - ďalšie miesta, na ktoré sa viažu pomocné proteíny (tzv. translačné faktory)
Najprv je rozoznávaná tRNA s iniciačnou aminokyselinou, ktorá vytvorí komplex s voľnou malou ribozomálnou podjednotkou a mRNA. Tohto rozpoznávania sa zúčastňujú aj pomocné translačné proteíny, tzv. iniciačné faktory. Tie okrem iného zabezpečia, že iniciačná aminokyselina vstúpi rovno do P miesta, kde sa nachádza iniciačný kodón AUG, tzn. že sa preskočí A miesto.
Genetický kód na mRNA sa číta po tripletoch, ktoré sú rozoznávané príslušnými tRNA s naviazanými aminokyselinami. Tieto tRNA vstupujú do A miesta, kde ich nasmeruje jeden z elongačných faktorov. Ak sa antikodón tRNA správne spáruje s kodónom na mRNA, peptidyltransferáza vytvorí peptidovú väzbu medzi novou aminokyselinou a rastúcim polypeptidovým reťazcom. Potom sa ribozóm posunie presne o jeden kodón pozdĺž mRNA (translokácia ribozómu), čo presunie deacylovanú tRNA do E miesta, polypeptid napojený na tRNA do P miesta a A miesto sa uvoľní.
Syntéza proteínu končí, ak ribozóm narazí na jeden z terminačných kodónov (UAA, UAG, UGA). K týmto kodónom nie sú komplementárne antikodóny žiadnych tRNA nesúcich aminokyselinu. Namiesto toho sú rozoznávané terminačnými faktormi, ktoré sa svojou 3D štruktúrou podobajú aminoacyl-tRNA (tzv. molekulárne mimikry). Ich účinkom dôjde k hydrolýze väzby medzi tRNA a polypeptidovým reťazcom a jeho uvoľneniu z ribozómu.
Podobne ako v prípade transkripcie, hotový translačný produkt podlieha zmenám, na ktorých konci je proteín schopný vykonávať svoju biologickú funkciu. To zahŕňa odštiepenie alebo modifikáciu iniciačnej aminokyseliny, chemickú modifikáciu ďalších aminokyselín, tvorbu disulfidických mostíkov medzi cysteínovými zvyškami, pripojenie cukorných zvyškov (glykoproteíny) alebo prostetických skupín (napr. hem).
U Escherichia coli prebieha syntéza polypeptidového reťazca rýchlosťou 10−20 aminokyselín za sekundu. Napodiv je tento proces veľmi presný, pretože frekvencia zaradenia nesprávnej aminokyseliny, často však podobného chemického zloženia, je 1 : 2000.
U baktérií nastupuje translácia často ešte pred ukončením transkripcie mRNA. Pritom najdôležitejší translačný signál predstavuje tzv. vedúca sekvencia (Shine-Dalgarnova sekvencia) 5'-AGGA-3' umiestnená ešte pred iniciačným kodónom na mRNA. Tá je komplementárna k sekvencii 3'-UCCU-5', ktorá sa nachádza na 3'-konci 16S-rRNA na malej ribozomálnej podjednotke. Prokaryotická mRNA tak už počas svojej syntézy môže byť pokrytá mnohými ribozómami, čo sa označuje pojmom polyzóm. Bezprostredné časové a priestorové prepojenie bakteriálnej transkripcie s transláciou využívajú niektoré regulovateľné transkripčné jednotky.
Typickým znakom bakteriálnej translácie je modifikácia iniciačnej aminokyseliny metionínu na formylmetionín. Pripájanie karbonylovej skupiny na NH₂-skupinu metionínu prebieha po naviazaní metionínu na iniciačnú transferovú RNA, s označením tRNAfMet. Takáto tRNAfMet s naviazaným formylmetionínom je rozoznávaná iniciačným faktorom IF2, čo zabezpečuje, že sa formylmetionín viaže na iniciačný kodón AUG a tvorí prvú aminokyselinu v polypeptidovom reťazci.
Po zaradení formylmetionínu často nastáva jeho modifikácia (odstránenie karbonylovej skupiny) a až v 50 % prípadov dochádza neskôr k odštiepeniu iniciačnej aminokyseliny. Všetky iniciačné faktory (IF1 až IF3) sa viažu len s voľnou malou ribozomálnou podjednotkou a po spojení ribozómu sa z nej uvoľňujú.
Elongácia translácie prebieha podľa všeobecného princípu popísaného vyššie. Čo sa však stane, ak sa vo vnútri čítacieho rámca objaví ďalší iniciačný kodón AUG? Nič zvláštne. Ten je rozoznávaný transferovou RNA s označením tRNAMet s naviazaným metionínom, ktorý vytvorí komplex s elongačným faktorom EF-Tu, a ten spôsobí, že tRNAMet vstúpi do A miesta podobne ako aj ostatné tRNA pri elongácii. Podobný princíp rozoznávania START kodónu a vnútorných AUG kodónov funguje aj u eukaryot.
Translokáciu ribozómu zabezpečuje elongačný faktor EF-G, ktorý hydrolyzuje GTP. U eukaryot je translácia oddelená od transkripcie časovo aj priestorovo (transkripcia v jadre, translácia v cytoplazme). Eukaryotickej mRNA chýba Shine-Dalgarnova sekvencia, ktorá je prítomná u prokaryot. Rozlišovacím prvkom je v tomto prípade post-transkripčne modifikovaný 5'-koniec mRNA, ktorý je rozoznávaný eukaryotickými iniciačnými faktormi.
Princíp tvorby komplexu mRNA, malej ribozómovej podjednotky a iniciačnej tRNA je rovnaký ako u prokaryot. Po jeho vytvorení dochádza k posúvaniu malej ribozomálnej podjednotky po reťazci mRNA až po iniciačný kodón AUG, kde dôjde k jeho rozoznaniu a spojeniu malej podjednotky ribozómu s veľkou za súčasného uvoľnenia iniciačných faktorov. Tento tzv. scanning model súvisí s tým, že AUG kodón je od 5'-konca mRNA značne vzdialený v porovnaní so vzdialenosťou medzi AUG kodónom a Shine-Dalgarnovou sekvenciou u prokaryot.
Translokácia ribozómu. Z E miesta vypadne pôvodná deacylovaná tRNA a jej miesto obsadí nová deacylovaná tRNA, peptidyl-tRNA sa posunie do P miesta a A miesto sa uvoľní. Translácia končí vyviazaním všetkých molekúl z mRNA.
Nukleové kyseliny a nukleotidy
Nukleové kyseliny sú biomakromolekulové látky zodpovedné za organizáciu a reprodukciu živej hmoty. Nukleové kyseliny sú polynukleotidy, ktorých stavebnou jednotkou sú nukleotidy. Nukleotid sa skladá z troch častí:
- zásaditá - heterocyklické dusíkaté purínové alebo pyrimidínové bázy,
- neutrálna - pentózy D-ribóza alebo 2-deoxy-D-ribóza,
- kyslá - kyselina trihydrogénfosforečná H₃PO₄.
Dusíkaté bázy sú heterocyklické zlúčeniny, kde heteroatómom je dusík. Nukleotid vzniká vzájomnou kondenzáciou všetkých troch zložiek, pričom sa uvoľňujú molekuly vody. Nukleotid je nukleozid esterifikovaný kyselinou trihydrogénfosforečnou. Nukleotidy a nukleozidy nie sú len stavebnou jednotkou nukleových kyselín. V bunkách môžu plniť rôzne funkcie. Zúčastňujú sa napríklad na prenose vodíka (NAD - nikotínamid-adenín-dinukleotid, FAD - flavín-adenín-dinukleotid), na prenose charakteristických skupín (napr. -NH₂), na biosyntéze bielkovín, sacharidov a lipidov.
Obzvlášť dôležitými nukleotidmi sú AMP (adenozín-monofosfát), ADP (adenozín-difosfát) a hlavne ATP (adenozín-trifosfát), ktoré sa zúčastňujú prenosu voľnej chemickej energie. V bunke plnia úlohu tzv. „univerzálneho energetického platidla”. ATP predstavuje primárny zdroj energie pre bunku. Energia sa uvoľňuje pri hydrolytickom štiepení molekuly kyseliny trihydrogénfosforečnej z nukleotidu ATP. Takto získaná energia je väčšia ako pri hydrolýze iných typov zlúčenín, preto ATP nazývame makroergickou zlúčeninou a makroergickú väzbu označujeme vlnovkou (nie je to nijako špeciálny typ väzby, je to proste obyčajná kovalentná väzba a vlnovkou zdôrazňujeme len jej energetický význam). cAMP (cyklický adenozín-monofosfát) je dôležitou signálnou molekulou alebo regulátorom génovej expresie.
Polynukleotid vzniká tak, že sa jednotlivé nukleotidy navzájom viažu intranukleotidovou esterovou väzbou pomocou H₃PO₄ medzi uhlíkmi C3 a C5 dvoch susedných ribóz (ribofuranóz).
DNA
Deoxyribonukleová kyselina sa skladá z týchto častí:
- dusíkaté bázy: purínové (adenín, guanín), pyrimidínové (tymín, cytozín),
- cukor: 2-deoxy-D-ribóza,
- kys. trihydrogénfosforečná.
Primárna štruktúra DNA je daná počtom a poradím, t.j. sekvenciou nukleotidov, ktoré sú v polynukleotidovom reťazci pospájané 3',5'-fosfodiesterovou väzbou. Relatívna molekulová hmotnosť takejto makromolekuly môže byť rádovo až 109 Da, čo predstavuje asi 200 tisíc stavebných jednotiek.
Primárna štruktúra DNA určuje poradie aminokyselín v bielkovinách. Sekvencia nukleotidov, ktorá obsahuje informáciu pre syntézu určitého produktu (najčastejšie je ním funkčná bielkovina), sa nazýva gén. Gén je taktiež základnou jednotkou dedičnosti z pohľadu genetiky. Molekulu DNA tvoria v skutočnosti dva polynukleotidové reťazce ležiace tesne nad sebou, pričom oproti A jedného vlákna sa vždy nachádza T na druhom vlákne, a oproti C sa vždy nachádza G. Toto párovanie sa označuje ako komplementarita dusíkatých báz. Umožňuje tvorbu presnej kópie molekuly DNA a zabezpečuje kontinuitu života.
Dva protiľahlé reťazce DNA sa "párujú" v dôsledku medzimolekulových interakcií v podobe vodíkových mostíkov, pričom medzi A-T sú 2 vodíkové mostíky a medzi C-G sú 3 vodíkové mostíky. Počet vodíkových mostíkov určuje silu tejto väzby, ktorá má význam pre mnohé deje súvisiace s metabolizmom DNA (napr. v oblastiach bohatých na A-T dochádza ľahšie k rozpletaniu reťazcov pri začiatku replikácie DNA).
J. WATSON a F. CRICK navrhli na základe špeciálnej röntgenovej (RTG) analýzy v roku 1953 priestorový model DNA (Nobelova cena, r. 1962), ktorým je vyjadrená jej sekundárna štruktúra. Sekundárnu štruktúru DNA tvoria dva polynukleotidové reťazce, ktoré sú antiparalelne spletené do pravotočivej dvojzávitnice, tzv. dvojitý α-helix. Táto štruktúra je pomerne stabilná.
Antiparalelná je taká konštelácia dvojzávitnice DNA, že oproti reťazcu, ktorý má fosfodiesterové väzby v smere 3'-5', je naložený druhý reťazec orientovaný v opačnom smere, tzn. 5'-3'.
Terciárna štruktúra DNA vzniká stočením dvojzávitnice v priestore do tzv. superhelixu.
RNA
Ribonukleová kyselina sa skladá z týchto častí:
- dusíkaté bázy: purínové (adenín, guanín), pyrimidínové (uracil, cytozín),
- cukor: D-ribóza,
- kys. trihydrogénfosforečná.
Primárna štruktúra RNA je v podstate rovnaká ako u DNA. Rozdiel je len v zložení nukleotidov, v ktorých namiesto deoxyribózy sa nachádza ribóza a z dusíkatých báz namiesto tymínu T sa nachádza uracil U. Párovanie báz pri tvorbe heteroduplexov alebo dvojzávitnicových molekúl je potom takéto: A-U a G-C.
Pojmom heteroduplex sa označuje dvojzávitnica tvorená kombináciou jednovláknových molekúl DNA-RNA, ktorá vzniká napríklad v priebehu transkripcie, kedy RNA-polymeráza pripája nukleotidy RNA na jednovláknovú matricu DNA. Treba zdôrazniť, že takýto heteroduplexný úsek DNA-RNA je len veľmi krátky a rýchlo sa pôsobením polymerázy rozpadá na DNA a predlžujúcu sa RNA.
Sekundárna a terciárna štruktúra RNA je oveľa rôznorodejšia ako v prípade DNA. Pri izolácii RNA z bunky dostaneme vzorku, ktorá obsahuje rôzne typy tejto nukleovej kyseliny. Dokonca aj molekula jedného typu RNA nemusí mať rovnakú priestorovú štruktúru. Veľká väčšina RNA je jednovláknová, a práve vďaka tomu sa môže stať, že pri ohybe vlákna dôjde k vzniku vnútromolekulových vodíkových mostíkov.
- Mediátorová RNA (mRNA) (angl. messenger = posol) - alebo informačná (iRNA), predstavuje 5-10 % obsahu RNA v bunke. Jej úlohou je prenos genetickej informácie z DNA do štruktúry bielkovín. Tvorí sa priamo na reťazci DNA na základe komplementarity báz, takže A z DNA sa prepíše ako U do RNA, G sa prepíše ako C, T ako A, a C ako G. Má jednovláknovú štruktúru a jej dĺžka závisí od množstva informácií (dĺžky génu), ktoré nesie. Trojice nukleotidov v mRNA nazývané kodóny predstavujú pri proteosyntéze informáciu pre zaradenie jednej aminokyseliny do polypeptidového reťazca.
- Ribozómová RNA (rRNA) tvorí základnú stavebnú zložku ribozómov a predstavuje až 90 % obsahu RNA v bunke. Priamo na ribozómoch dochádza k vlastnej syntéze proteínov (proteosyntéze). Časti makromolekuly rRNA môžu mať tvar dvojitej závitnice. Ribozómové RNA sa najčastejšie charakterizujú sedimentačným koeficientom S.
- Transferové RNA (tRNA) majú špecifickú sekundárnu a terciárnu štruktúru, ktorá je daná bohatým vnútromolekulovým párovaním báz. Transferová RNA zabezpečuje prenos (transfer) aktivovaných aminokyselín na miesto tvorby polypeptidového reťazca - do ribozómu. Špecifickosť tRNA pre zaradenie danej aminokyseliny určuje trojica nukleotidov v strednej časti molekuly - antikodón, ktorým tRNA rozoznáva kodóny na mRNA. Súhrn pravidiel, podľa ktorých príslušný kodón na mRNA vedie k zaradeniu určitej aminokyseliny do polypeptidového reťazca, sa nazýva genetický kód. Transferové RNA nemajú veľkú molekulovú hmotnosť. Obsahujú okolo 70 až 80 nukleotidov.
tags: #trojica #nukleotidov #ktora #urcuje #zaradenie #aminokyseliny